


Нові дані з систематики та еволюції птахів, які були отримані в рамках проекту Genome 10k
В систематиці сучасних птахів є багато спірних та невирішених питань. Більшість їх груп беруть свій початок з раннього Кайнозою, близько 60 мільйонів років тому, коли відбулась швидка адаптивна радіація. Такий бурхливий розвиток ускладнив розв’язання систематичних відносин. Кожен раз з появою нових методів систематика птахів суттєво переглядалась. Нещодавно був зроблений ще один прорив в цьому напрямку: в рамках великого міжнародного проекту було проведено прочитання повних геномних послідовностей представників майже всіх сучасних рядів птахів. Використання нових методів обробки даних дозволило уточнити родинні зв’язки птахів між собою та деякі особливості еволюції цієї групи.

Всі сучасні птахи відносяться до групи Neornithes, або віялохвостих птахів. Для них характерні скорочений хвіст, в якому всі хвостові пера кріпляться віялом до пігостилю – кістки, сформованої кількома зрощеними хребцями, відсутність зубів та інші ознаки. Ця група виникла близько 120 мільйонів років тому, на початку крейдового періоду. Згідно сучасним уявленням найближчими родичами Neornithes були зубаті гесперорніси.
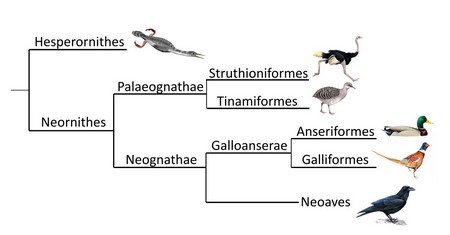
Філогенетичні зв’язки сучасних птахів (Neornithes)
Велике вимирання на рубежі крейдового та палеогенового періодів не обійшло і групу сучасних птахів. Багато представників цього підкласу вимерли, не залишивши сучасних нащадків. Після вимирання відбулась дуже швидка адаптивна радіація птахів – поява нових життєвих форм, які зайняли вільні екологічні ніші. Більшість рядів сучасних птахів виникли саме в цей період. Швидке виникнення нових груп призвело до появи багатьох конвергенцій в різних систематичних групах та ускладнило встановлення еволюційних взаємовідносин між ними.
В систематиці птахів завжди було багато спірних питань. Через особливості будови скелету залишки птахів погано зберігаються, тому для них завжди була характерною неповнота палеонтологічного літопису. А це основний матеріал для встановлення еволюційних відносин між видами. Зараз немає єдиного погляду на те, скільки існує рядів та родин птахів та які групи до них відносяться. Ці суперечності важливі не лише для купки систематиків, яким кортить створити правильні та красиві списки видів, згрупованих в роди, родини та ряди. Принципи сучасної систематики живих організмів базуються на філогенетичному підході, тобто різні види об’єднують на основі їх еволюційної спорідненості між собою.
На ранніх етапах систематики птахів використовували анатомічні та зовнішні морфологічні ознаки. Пізніше в систематичні дослідження включали біохімічні, фізіологічні та поведінкові ознаки, а для уточнення спорідненості видів між собою ще дані палеонтології та зоогеографії. В середині минулого сторіччя найбільш популярною була система Ветмора, яка виділяла 27 сучасних рядів, в тому числі пінгвінів в складі окремого надряду. Ця система була прийнята в відомому радянському виданні «Жизнь животных» та багатьох інших книжках. Зараз більш розповсюдженою та загальноприйнятою в Європі є система Клементса, яка виділяє два підкласи – безкилеві (Paleognathae) та кілегруді (Neognathae), в складі яких також 27 сучасних рядів. В останні роки дослідження систематики та філогенії птахів ведуть з використанням методів молекулярної біології та, зокрема, вивчення послідовностей ДНК. Першим проривом в цьому напрямку була відома класифікація Сіблі-Алквіста, розроблена в 1970-80 х роках. За основу автори взяли метод гібридизації ДНК, який дозволяє оцінювати ступень відмінності молекул ДНК різних організмів між собою, але не дає даних про послідовність нуклеотидів. Отримані ними результати істотно відрізнялись від попередніх досліджень: одні ряди були об’єднані з іншими та збільшені, інші розбиті на більш дрібні, змінено систематичне положення деяких родин. Попри те, що частина коректив стали загальноприйнятими, ця система викликала багато критики та зараз є більш популярною в Сполучених Штатах Америки. Паралельно з цим існують кілька систем, які намагаються поєднати нові молекулярні дані з результатами інших досліджень.
Найбільш інформативним молекулярним методом є вивчення послідовностей, або прочитання молекул-носіїв спадкової інформації, ДНК та РНК. Воно дозволяє знаходити відмінності як на рівні особин, так і на рівні різних рядів, зіставляти їх з біологічними властивостями організмів та класифікувати. Але з огляду на те, що розмір генома хребетних складає мільярди пар нуклеотидних основ, ці методи потребують значного розвитку технологій та суттєвих зусиль для обробки. До теперішнього часу у птахів досліджували лише послідовності окремих генів або груп генів, а не всього ядерного геному.
Протягом останніх 15 років розвиток технологій забезпечив скорочення вартості прочитання одного генома з сотень мільйонів до тисяч доларів. Це призвело до збільшення доступності досліджень послідовностей ДНК та збільшення кількості видів, для яких був прочитаний весь геном. В 2008 році було опубліковано масштабне дослідження, основане на прочитанні великих частин геному деяких видів птахів, але не цілих ядерних геномів. Логічним продовженням цього стали роботи в рамках проекту Genome 10k започаткованого у 2009 р. Його ціллю було прочитання повних геном них послідовностей 10 тисяч видів хребетних тварин. Одним з дочірніх проектів став Avian Phylogenomics Project, який відповідає за дослідження птахів. В цьому проекті працюють сотні дослідників з більш ніж 200 країн. З огляду на постійний розвиток молекулярних методів свою ціль вони планують досягти протягом наступного десятиліття, але перший етап робіт був завершений вже у минулому році.
До початку роботи повні геноми були прочитані лише у трьох видів птахів: курки, індички та зебрової амадини. Дослідники обрали по одному представнику кожного сучасного ряду птахів, окрім деяких представників палеогнат (тільки африканський страус та тинаму). У горобцеподібних, папуг, денних хижих птахів, пінгвінів, куроподібних було обрано по кілька представників для аналізу так, щоб сполучались найменш споріднені види. Таким чином, додатково прочитали геноми 45 видів. В результаті були отримані настільки великі об’єми даних, що для їх аналізу знадобилась суттєва модифікація існуючого програмного забезпечення та розробка нових алгоритмів аналізу даних. Навіть після цього обробка даних з використанням дев’яти центрів суперкомп’ютерів зайняла кілька років. Роботи за проектом стартували в 2010 р., але тільки в 2014 р. була завершена перша серія аналізу даних.
Головними питаннями, які стояли перед дослідниками, було розв’язання спірних моментів в еволюції птахів, у тому числі достовірність виділених попередніми дослідниками окремих груп. Особливу увагу приділяли групам із нез’ясованим систематичним положенням: сивкоподібним, птахам-мишам, совам та іншим. За допомогою молекулярних методів встановлювали строки появи певних груп та розвиток окремих рис серед них.
Математична обробка отриманих даних дозволила побудувати кладограму (філогенетичне древо), на якій відображені родинні зв’язки між рядами сучасних птахів. Дослідники відмічають, що різні способи розрахунку давали близькі за своїм складом кладограми, а ступінь достовірності виявлених зв’язків між більшістю груп була дуже високою.
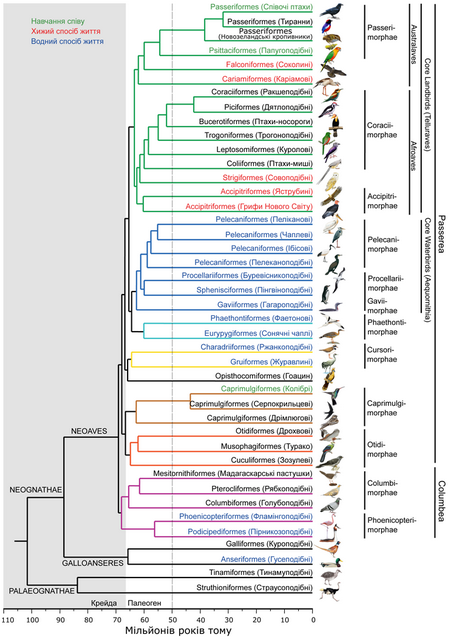
Філогенія сучасних птахів за результатами дослідження. Довжина гілок відповідає періодам розходження близьких груп, внизу наведена часова шкала у мільйонах років тому. Синім кольором виділені назви таксонів, пристосованих до водного способу життя, червоним – хижаків, зеленим – груп, серед яких зустрічається навчання співу.За матеріалами Jarvis et al., 2014, зі змінами.
По-перше ці результати, отримані на принципово більших обсягах даних, підтвердили достовірність виділених раніше груп: Palaeognathae (страуси, тінаму та інші), Galloanseres (дуже близькі один до одного ряди гусеподібні та куроподібні), Neoaves (всі інші окрім попередніх двох груп). Остання група ділиться на дві великі гілки: Columbea (голубоподібні, фламінгоподібні, пірникозоподібні)та Passerea (більшість інших груп), між якими спостерігаються багато конвергенцій.
В одну групу були об’єднані такі складні систематичні групи, як дрохви, турако та зозулі, що доводить їх спільне походження та неспорідненість дрохв з журавлями та пастушками, а зозуль з гоацином.
Гоацин взагалі є дуже складним видом для систематиків. У різних системах він «кочував» по різних рядах. Навіть в рамках проекту Genome 10k не вдалося з певністю встановити його родинні зв’язки, але достатньо точно відомо, що гоацини – це одна з найдавніших груп серед Neoaves, яка виникла на початку Кайнозою.

Гоацин – один з тих видів птахів, визначення систематичного положення якого завжди викликало надзвичайні труднощі. Предки гоацину досить давно відокремились від інших птахів.
Систематичне положення ще однієї складної групи, птахів-мишей, було уточнено: вони віднесені до однієї клади з ракшеподібними та трогоноподібними.
Достовірність виділення ряду філогенетичних груп попередніми дослідниками не була підтверджена на основі аналізу повних геномів. Так, в одній з попередніх робіт було запропоновано виділити дві великі групи Metaves та Coronaves (тільки на основі аналізу гену β-фібріногену), але це не знайшло підтвердження в роботі. Але в об’єднанні Core landbirds були виділені дві великі групи Australaves та Afroaves, серед яких зустрічається багато конвергентно схожих форм.
Було виявлено багато цікавого в частині спорідненості таксонів, до складу яких входять ззовні несхожі між собою птахи. Так, грифи Старого Світу виявились окремою групою від яструбиних, але спорідненість їх з лелекоподібними, про яку говорили попередні дослідники, доведена не була. Фаетони проявили себе як не споріднені з пеліканоподібними, до яких їх відносили.


Попри зовнішню несхожість пірникози та фламінго виявились близькими родичами.
В традиційних системах існують кілька дуже різнорідних рядів, які містять слабо споріднені групи, що добре відрізняються одна від одної. Перша з них – це ракшеподібні. Тут виявилось, що птахи-носороги та куроли по суті заслуговують на виділення в окремі ряди, а дятлоподібні стоять до «традиційних» ракшеподібних значно ближче. Журавлеподібні власне взагалі не мають відношення до традиційно віднесених до них каріам (ближче до соколів), дрохв (до зозулеподібних) та сонячних чапель (до фаетонів).
Сучасні аналітичні методи дозволяють не тільки встановлювати ступінь спорідненості різних груп між собою, але й приблизно визначати строки їх виникнення. Більшість отриманих в роботі датувань була цілком співставна з наявними палеонтологічними даними.
Так, розходження між еволюційними гілками неогнат та палеогнат відбулось приблизно 100 мільйонів років тому. Розділення предків страусів та тинамових – близько 84 мільйонів років тому. Дві основні групи серед неогнат – Galloanseres та Neoaves – розійшлись 88 мільйонів років тому, задовго до початку радіації, яка призвела до виникнення більшості сучасних рядів. Отже, гусеподібні та куроподібні є досить давніми групами, відокремленими від більшості сучасних птахів.

Vegavis – вимерлі представники гусеподібних (клада Galloanseres) з крейдового періоду, описані в 2005 році. Ця знахідка стала доказом того, що деякі сучасні групи птахів сформувались ще в мезозої. Джерело – Scientific American.
Швидка радіація (розходження) серед Neoaves дійсно відбулась в досить короткий в еволюційному сенсі час: більшість сучасних рядів птахів з’явились за період яких-небудь 6,5 мільйонів років. Але поява нових сучасних груп розпочалась не після, а до крейдового масового вимирання 65 мільйонів років тому: ще тоді з’явились кілька великих груп, еволюція в яких потім йшла паралельними шляхами.
Дуже прикметними є чисельні конвергенції, які виникли в різних еволюційних гілках птахів. Наприклад, пристосування до водного способу життя зустрічаються в трьох великих групах: Galloanseres (гусеподібні), Columbea (пірникозоподібні та фламінгоподібні) та Core waterbirds в Passerea (всі інші групи). При цьому відомо, що спільні предки цих груп були наземними тваринами.
Важливою особливістю птахів, яку вони поділяють лише з деякими групами ссавців, є здатність навчатися співу. Такі ознака є лише у кількох груп: колібрі, папуг та співочих птахів. З огляду на те, що еволюційні лінії колібрі розійшлись з іншими ще навіть до крейдового вимирання, а серед споріднених з папугами горобцеподібних є підряди тиранни та новозеландські кропивники, для яких навчання співу не властиве, можна зробити висновок, що ця ознака в історії птахів незалежно виникала два або три рази. До речі, спеціальне дослідження в рамках цього проекту довело існування багатьох конвергенцій між генетичними та нейрофізіологічними механізмами виникнення навчання співу у птахів та навчання мови у людини. Це свідчить, що діапазон потенціальних можливостей виникнення такої поведінки у хребетних тварин досить обмежений.
В групі Core landbirds птахи з хижим способом життя виявились розкидані по всім гілкам, при чому для більш давніх за походженням груп (соколині, яструбині, сови) хижий спосіб життя був більш властивим, ніж для більш молодих груп (горобцеподібні, дятлоподібні, птахи-носороги). Це наводить на думку, що спільні предки цієї найбільш багатої на сучасні види гілки птахів виглядали як середні за розмірами неспеціалізовані хижаки, що мешкали на зорі палеогенового періоду, невдовзі після вимирання динозаврів. Автори підкреслюють, що навіть викопні предки сучасних птахів-мишей та куролів вели більш хижий спосіб життя, ніж їх сучасні нащадки, а каріами є найближчими нащадками славнозвісних фороракосових птахів.
Аналізуючи чисельні конвергенції між різними групами, дослідники приходять до цікавого висновку, що в ранні періоди еволюції сучасних груп птахів могли мати місце неодноразові випадки гібридизації між представниками різних філогенетичних гілок, які тоді ще не розійшлись надто далеко. Це могло призвести до поширення однакових груп генів в різних групах птахів. Розрахунки показали, що головним механізмом такої швидкої радіації птахів на рубежі мезозою та кайнозою було зниження вірогідності вимирання таксонів, що виникали, тоді як темпи появи нових таксонів залишалися традиційними.
Немалозначним фактом, який дозволив в рамках даного проекту проаналізувати геноми птахів швидше за інших хребетних тварин було те, що вони є порівняно дуже маленькими. Для прикладу, якщо середній розмір пташиного геному складає 1 мільярд пар основ, у ссавців – 3 мільярди, а у амфібій від 6 до 9. З чим це пов’язано?
Геном повинен мати закодованими всі гени, що необхідні для нормального розвитку та життєдіяльності організму. Але окрім кодуючих послідовностей (екзонів) в ньому присутні досить великі ділянки, які не кодують білки (інтрони та багато інших). Частка некодуючих елементів в геномі людини складає 50%, тоді як у птахів – лише 5-10%. До того ж у деяких організмів увесь геном може існувати в клітинах у вигляді декількох копій. Аналіз ядерного геному птахів доводить, що зменшення розміру їх геному відбулось насамперед завдяки скороченню числа цих некодуючих послідовностей. Тобто птахи на певному етапі свого розвитку пройшли потужний відбір на скорочення розміру геному. Дослідники пов’язують це з необхідністю швидкої генної регуляції та зменшення загального об’єму тіла як пристосування до польоту.
На перший погляд економія на якихось шматках молекул ДНК здається беззмістовною, але треба мати на увазі, що збереження та функціонування ДНК в клітині потребує багато додаткових біохімічних механізмів, які також впливають на розмір клітини та її енергоспоживання. Це, в свою чергу, впливає на швидкість поділу клітини, а через це, на швидкість росту організму (докладніше про те, які наслідки несе збільшення розміру геному, можна прочитати тут - http://elementy.ru/news/432513). Доказом цих закономірностей є вивчення геномів соколів в рамках даної роботи, яке свідчить, що ці надзвичайно активні птахи пройшли ще більш жорсткий відбір на зменшення розміру геному.
Ще однією важливою ознакою птахів є відсутність зубів в щелепах. Ця ознака не дуже розповсюджена серед хребетних тварин:зубів не має у черепах, які втратили зуби значно раніше, ніж птахи, та частково у деяких ссавців (кити, неповнозубі та деякі інші).
Головним питанням беззубості птахів є те, чи втрата зубів відбулась у загального предка всіх сучасних птахів, або у предків різних сучасних груп незалежно. За формування зубів відповідні шість генів, які є спільними для всіх хребетних тварин. Сучасні методи дозволяють встановити, які з цих генів були деактивовані у беззубих форм, та коли приблизно це відбулось.
В процесі порівняння геномів виявилося, що залишки цих генів є в усіх беззубих форм хребетних. У різних груп ссавців до втрати зубів призвели різні мутації, що свідчить про незалежність процесів у різних групах. У всіх сучасних груп птахів залишки генів, що відповідали за формування зубів, мають схожі мутації, що говорить про втрату зубів ще їх далекими спільними предками. Аналіз показав, що інактивація генів, що відповідають за втрату емалі, відбулась приблизно 116 мільйонів років тому, а дентину – 101 мільйон років тому. Одночасно з цим формувались гени, відповідальні за формування рамфотеки (рогового чохла дзьобу). Дослідження свідчили, що втрата зубів починалась з переднього краю щелеп, там саме починав формуватись дзьоб, і після цього процес зсувався назад, охоплюючи всю щелепу.
Цікаво, що серед птахів в широкому сенсі втрата зубів відбувалась неодноразово. Так, в крейдовому періоді поруч з предками сучасних птахів, неорнісами (частина з яких була зубатою, а частина – беззубою), існували енанціорніси, які незалежно втратили зуби, та ще деякі зубаті форми, нащадки яких вимерли наприкінці періоду. Автори доходять висновку, що «у птахів існувала еволюційна схильність до втрати зубів».
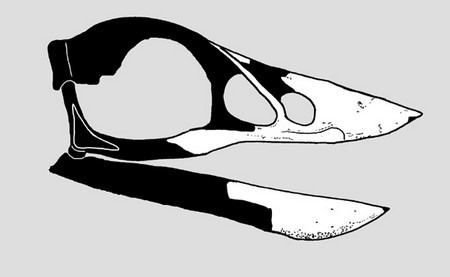
Череп Gobypterix minuta – одного з небагатьох енанціорнісів, який втратив зуби. Мешкав у другій половині крейдового періоду.

Реконструкція Gansus – одного з найбільш давніх представників віялохвостих птахів, що мешкав в крейдовому періоді близько 110 мільйонів років тому. Ці птахи мали зуби.
Безумовно, поточне дослідження стало важливим етапом у вивченні еволюційної історії птахів. Нові методи призвели до багаторазового збільшення обсягів зібраної інформації та підштовхнули до розробки нових способів аналізу даних. Але говорити про повне з’ясування питань систематики птахів ще зарано. З високим ступенем достовірності були підтверджені деякі останні ідеї щодо загальної еволюції птахів, окреслені загальні риси процесів. Але потрібна ще велика робота по розв’язанню питань еволюції менших за рангом таксонів. Значна частина проаналізованих груп самі по собі є дуже різноманітними, і включення більшого числа видів допоможе краще зрозуміти цей заплутаний ранній період еволюції птахів. Самі автори кажуть, що поступ в основному можливий у частині розробки більш вдосконалених методів аналізу геномних даних та у включенні все більшого числа видів до аналізу.
Дані молекулярних методів все ще мають багато протиріч з більш «традиційною» систематикою. Схоже, що для синтезу даних цих двох підходів потрібен деякий час, коли відбуватиметься накопичення нових матеріалів, розробка ефективних методів їх аналізу та узгодження з результатами інших досліджень.
Загалом можна сказати що відповіді на багато питань еволюції птахів криються в тому короткому та бурхливому періоді в 6 мільйонів років, підчас якого в основному і було закладене те різноманіття, яке ми спостерігаємо зараз.
За матеріалами:

Всі сучасні птахи відносяться до групи Neornithes, або віялохвостих птахів. Для них характерні скорочений хвіст, в якому всі хвостові пера кріпляться віялом до пігостилю – кістки, сформованої кількома зрощеними хребцями, відсутність зубів та інші ознаки. Ця група виникла близько 120 мільйонів років тому, на початку крейдового періоду. Згідно сучасним уявленням найближчими родичами Neornithes були зубаті гесперорніси.
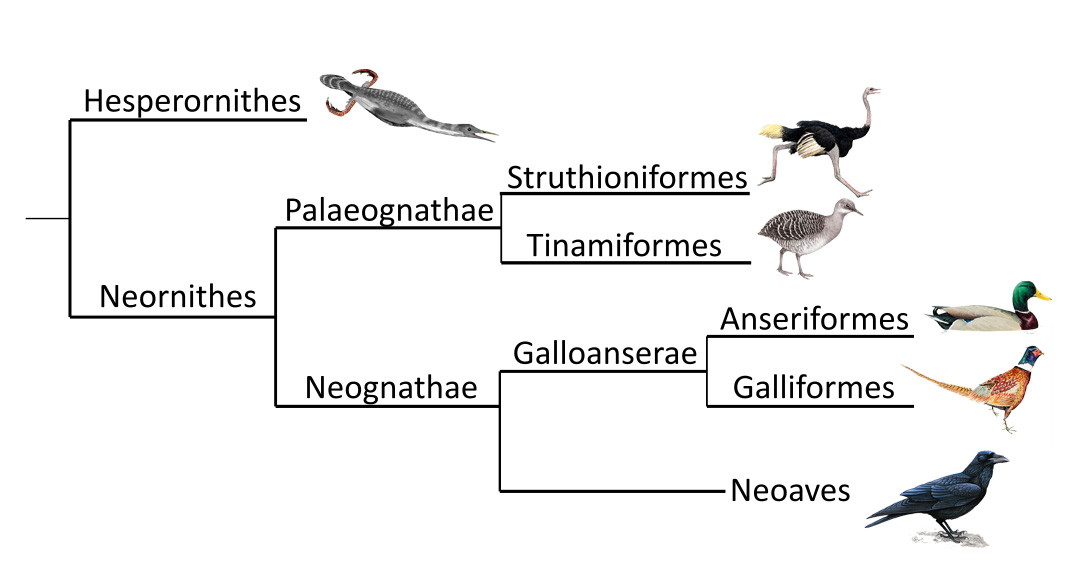
Філогенетичні зв’язки сучасних птахів (Neornithes)
Велике вимирання на рубежі крейдового та палеогенового періодів не обійшло і групу сучасних птахів. Багато представників цього підкласу вимерли, не залишивши сучасних нащадків. Після вимирання відбулась дуже швидка адаптивна радіація птахів – поява нових життєвих форм, які зайняли вільні екологічні ніші. Більшість рядів сучасних птахів виникли саме в цей період. Швидке виникнення нових груп призвело до появи багатьох конвергенцій в різних систематичних групах та ускладнило встановлення еволюційних взаємовідносин між ними.
В систематиці птахів завжди було багато спірних питань. Через особливості будови скелету залишки птахів погано зберігаються, тому для них завжди була характерною неповнота палеонтологічного літопису. А це основний матеріал для встановлення еволюційних відносин між видами. Зараз немає єдиного погляду на те, скільки існує рядів та родин птахів та які групи до них відносяться. Ці суперечності важливі не лише для купки систематиків, яким кортить створити правильні та красиві списки видів, згрупованих в роди, родини та ряди. Принципи сучасної систематики живих організмів базуються на філогенетичному підході, тобто різні види об’єднують на основі їх еволюційної спорідненості між собою.
На ранніх етапах систематики птахів використовували анатомічні та зовнішні морфологічні ознаки. Пізніше в систематичні дослідження включали біохімічні, фізіологічні та поведінкові ознаки, а для уточнення спорідненості видів між собою ще дані палеонтології та зоогеографії. В середині минулого сторіччя найбільш популярною була система Ветмора, яка виділяла 27 сучасних рядів, в тому числі пінгвінів в складі окремого надряду. Ця система була прийнята в відомому радянському виданні «Жизнь животных» та багатьох інших книжках. Зараз більш розповсюдженою та загальноприйнятою в Європі є система Клементса, яка виділяє два підкласи – безкилеві (Paleognathae) та кілегруді (Neognathae), в складі яких також 27 сучасних рядів. В останні роки дослідження систематики та філогенії птахів ведуть з використанням методів молекулярної біології та, зокрема, вивчення послідовностей ДНК. Першим проривом в цьому напрямку була відома класифікація Сіблі-Алквіста, розроблена в 1970-80 х роках. За основу автори взяли метод гібридизації ДНК, який дозволяє оцінювати ступень відмінності молекул ДНК різних організмів між собою, але не дає даних про послідовність нуклеотидів. Отримані ними результати істотно відрізнялись від попередніх досліджень: одні ряди були об’єднані з іншими та збільшені, інші розбиті на більш дрібні, змінено систематичне положення деяких родин. Попри те, що частина коректив стали загальноприйнятими, ця система викликала багато критики та зараз є більш популярною в Сполучених Штатах Америки. Паралельно з цим існують кілька систем, які намагаються поєднати нові молекулярні дані з результатами інших досліджень.
Найбільш інформативним молекулярним методом є вивчення послідовностей, або прочитання молекул-носіїв спадкової інформації, ДНК та РНК. Воно дозволяє знаходити відмінності як на рівні особин, так і на рівні різних рядів, зіставляти їх з біологічними властивостями організмів та класифікувати. Але з огляду на те, що розмір генома хребетних складає мільярди пар нуклеотидних основ, ці методи потребують значного розвитку технологій та суттєвих зусиль для обробки. До теперішнього часу у птахів досліджували лише послідовності окремих генів або груп генів, а не всього ядерного геному.
Протягом останніх 15 років розвиток технологій забезпечив скорочення вартості прочитання одного генома з сотень мільйонів до тисяч доларів. Це призвело до збільшення доступності досліджень послідовностей ДНК та збільшення кількості видів, для яких був прочитаний весь геном. В 2008 році було опубліковано масштабне дослідження, основане на прочитанні великих частин геному деяких видів птахів, але не цілих ядерних геномів. Логічним продовженням цього стали роботи в рамках проекту Genome 10k започаткованого у 2009 р. Його ціллю було прочитання повних геном них послідовностей 10 тисяч видів хребетних тварин. Одним з дочірніх проектів став Avian Phylogenomics Project, який відповідає за дослідження птахів. В цьому проекті працюють сотні дослідників з більш ніж 200 країн. З огляду на постійний розвиток молекулярних методів свою ціль вони планують досягти протягом наступного десятиліття, але перший етап робіт був завершений вже у минулому році.
До початку роботи повні геноми були прочитані лише у трьох видів птахів: курки, індички та зебрової амадини. Дослідники обрали по одному представнику кожного сучасного ряду птахів, окрім деяких представників палеогнат (тільки африканський страус та тинаму). У горобцеподібних, папуг, денних хижих птахів, пінгвінів, куроподібних було обрано по кілька представників для аналізу так, щоб сполучались найменш споріднені види. Таким чином, додатково прочитали геноми 45 видів. В результаті були отримані настільки великі об’єми даних, що для їх аналізу знадобилась суттєва модифікація існуючого програмного забезпечення та розробка нових алгоритмів аналізу даних. Навіть після цього обробка даних з використанням дев’яти центрів суперкомп’ютерів зайняла кілька років. Роботи за проектом стартували в 2010 р., але тільки в 2014 р. була завершена перша серія аналізу даних.
Головними питаннями, які стояли перед дослідниками, було розв’язання спірних моментів в еволюції птахів, у тому числі достовірність виділених попередніми дослідниками окремих груп. Особливу увагу приділяли групам із нез’ясованим систематичним положенням: сивкоподібним, птахам-мишам, совам та іншим. За допомогою молекулярних методів встановлювали строки появи певних груп та розвиток окремих рис серед них.
Філогенетичні результати
Математична обробка отриманих даних дозволила побудувати кладограму (філогенетичне древо), на якій відображені родинні зв’язки між рядами сучасних птахів. Дослідники відмічають, що різні способи розрахунку давали близькі за своїм складом кладограми, а ступінь достовірності виявлених зв’язків між більшістю груп була дуже високою.
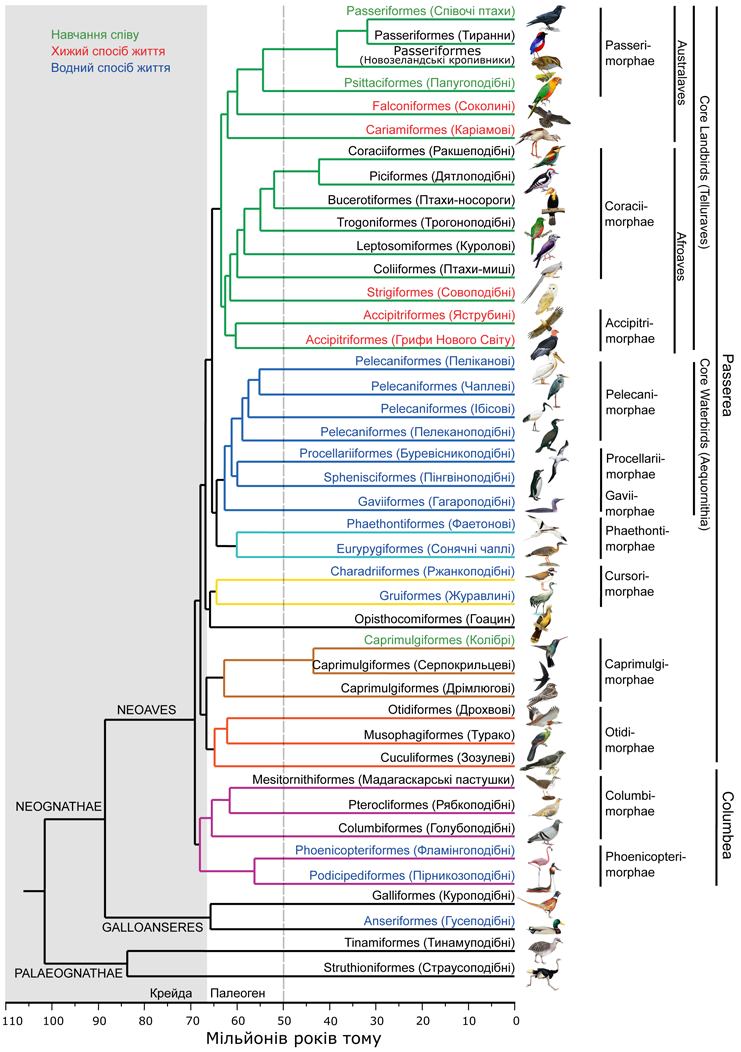
Філогенія сучасних птахів за результатами дослідження. Довжина гілок відповідає періодам розходження близьких груп, внизу наведена часова шкала у мільйонах років тому. Синім кольором виділені назви таксонів, пристосованих до водного способу життя, червоним – хижаків, зеленим – груп, серед яких зустрічається навчання співу.За матеріалами Jarvis et al., 2014, зі змінами.
По-перше ці результати, отримані на принципово більших обсягах даних, підтвердили достовірність виділених раніше груп: Palaeognathae (страуси, тінаму та інші), Galloanseres (дуже близькі один до одного ряди гусеподібні та куроподібні), Neoaves (всі інші окрім попередніх двох груп). Остання група ділиться на дві великі гілки: Columbea (голубоподібні, фламінгоподібні, пірникозоподібні)та Passerea (більшість інших груп), між якими спостерігаються багато конвергенцій.
В одну групу були об’єднані такі складні систематичні групи, як дрохви, турако та зозулі, що доводить їх спільне походження та неспорідненість дрохв з журавлями та пастушками, а зозуль з гоацином.
Гоацин взагалі є дуже складним видом для систематиків. У різних системах він «кочував» по різних рядах. Навіть в рамках проекту Genome 10k не вдалося з певністю встановити його родинні зв’язки, але достатньо точно відомо, що гоацини – це одна з найдавніших груп серед Neoaves, яка виникла на початку Кайнозою.

Гоацин – один з тих видів птахів, визначення систематичного положення якого завжди викликало надзвичайні труднощі. Предки гоацину досить давно відокремились від інших птахів.
Систематичне положення ще однієї складної групи, птахів-мишей, було уточнено: вони віднесені до однієї клади з ракшеподібними та трогоноподібними.
Достовірність виділення ряду філогенетичних груп попередніми дослідниками не була підтверджена на основі аналізу повних геномів. Так, в одній з попередніх робіт було запропоновано виділити дві великі групи Metaves та Coronaves (тільки на основі аналізу гену β-фібріногену), але це не знайшло підтвердження в роботі. Але в об’єднанні Core landbirds були виділені дві великі групи Australaves та Afroaves, серед яких зустрічається багато конвергентно схожих форм.
Було виявлено багато цікавого в частині спорідненості таксонів, до складу яких входять ззовні несхожі між собою птахи. Так, грифи Старого Світу виявились окремою групою від яструбиних, але спорідненість їх з лелекоподібними, про яку говорили попередні дослідники, доведена не була. Фаетони проявили себе як не споріднені з пеліканоподібними, до яких їх відносили.


Попри зовнішню несхожість пірникози та фламінго виявились близькими родичами.
В традиційних системах існують кілька дуже різнорідних рядів, які містять слабо споріднені групи, що добре відрізняються одна від одної. Перша з них – це ракшеподібні. Тут виявилось, що птахи-носороги та куроли по суті заслуговують на виділення в окремі ряди, а дятлоподібні стоять до «традиційних» ракшеподібних значно ближче. Журавлеподібні власне взагалі не мають відношення до традиційно віднесених до них каріам (ближче до соколів), дрохв (до зозулеподібних) та сонячних чапель (до фаетонів).
Подробиці ранньої еволюції птахів
Сучасні аналітичні методи дозволяють не тільки встановлювати ступінь спорідненості різних груп між собою, але й приблизно визначати строки їх виникнення. Більшість отриманих в роботі датувань була цілком співставна з наявними палеонтологічними даними.
Так, розходження між еволюційними гілками неогнат та палеогнат відбулось приблизно 100 мільйонів років тому. Розділення предків страусів та тинамових – близько 84 мільйонів років тому. Дві основні групи серед неогнат – Galloanseres та Neoaves – розійшлись 88 мільйонів років тому, задовго до початку радіації, яка призвела до виникнення більшості сучасних рядів. Отже, гусеподібні та куроподібні є досить давніми групами, відокремленими від більшості сучасних птахів.

Vegavis – вимерлі представники гусеподібних (клада Galloanseres) з крейдового періоду, описані в 2005 році. Ця знахідка стала доказом того, що деякі сучасні групи птахів сформувались ще в мезозої. Джерело – Scientific American.
Швидка радіація (розходження) серед Neoaves дійсно відбулась в досить короткий в еволюційному сенсі час: більшість сучасних рядів птахів з’явились за період яких-небудь 6,5 мільйонів років. Але поява нових сучасних груп розпочалась не після, а до крейдового масового вимирання 65 мільйонів років тому: ще тоді з’явились кілька великих груп, еволюція в яких потім йшла паралельними шляхами.
Дуже прикметними є чисельні конвергенції, які виникли в різних еволюційних гілках птахів. Наприклад, пристосування до водного способу життя зустрічаються в трьох великих групах: Galloanseres (гусеподібні), Columbea (пірникозоподібні та фламінгоподібні) та Core waterbirds в Passerea (всі інші групи). При цьому відомо, що спільні предки цих груп були наземними тваринами.
Важливою особливістю птахів, яку вони поділяють лише з деякими групами ссавців, є здатність навчатися співу. Такі ознака є лише у кількох груп: колібрі, папуг та співочих птахів. З огляду на те, що еволюційні лінії колібрі розійшлись з іншими ще навіть до крейдового вимирання, а серед споріднених з папугами горобцеподібних є підряди тиранни та новозеландські кропивники, для яких навчання співу не властиве, можна зробити висновок, що ця ознака в історії птахів незалежно виникала два або три рази. До речі, спеціальне дослідження в рамках цього проекту довело існування багатьох конвергенцій між генетичними та нейрофізіологічними механізмами виникнення навчання співу у птахів та навчання мови у людини. Це свідчить, що діапазон потенціальних можливостей виникнення такої поведінки у хребетних тварин досить обмежений.
В групі Core landbirds птахи з хижим способом життя виявились розкидані по всім гілкам, при чому для більш давніх за походженням груп (соколині, яструбині, сови) хижий спосіб життя був більш властивим, ніж для більш молодих груп (горобцеподібні, дятлоподібні, птахи-носороги). Це наводить на думку, що спільні предки цієї найбільш багатої на сучасні види гілки птахів виглядали як середні за розмірами неспеціалізовані хижаки, що мешкали на зорі палеогенового періоду, невдовзі після вимирання динозаврів. Автори підкреслюють, що навіть викопні предки сучасних птахів-мишей та куролів вели більш хижий спосіб життя, ніж їх сучасні нащадки, а каріами є найближчими нащадками славнозвісних фороракосових птахів.
Аналізуючи чисельні конвергенції між різними групами, дослідники приходять до цікавого висновку, що в ранні періоди еволюції сучасних груп птахів могли мати місце неодноразові випадки гібридизації між представниками різних філогенетичних гілок, які тоді ще не розійшлись надто далеко. Це могло призвести до поширення однакових груп генів в різних групах птахів. Розрахунки показали, що головним механізмом такої швидкої радіації птахів на рубежі мезозою та кайнозою було зниження вірогідності вимирання таксонів, що виникали, тоді як темпи появи нових таксонів залишалися традиційними.
Розмір геному
Немалозначним фактом, який дозволив в рамках даного проекту проаналізувати геноми птахів швидше за інших хребетних тварин було те, що вони є порівняно дуже маленькими. Для прикладу, якщо середній розмір пташиного геному складає 1 мільярд пар основ, у ссавців – 3 мільярди, а у амфібій від 6 до 9. З чим це пов’язано?
Геном повинен мати закодованими всі гени, що необхідні для нормального розвитку та життєдіяльності організму. Але окрім кодуючих послідовностей (екзонів) в ньому присутні досить великі ділянки, які не кодують білки (інтрони та багато інших). Частка некодуючих елементів в геномі людини складає 50%, тоді як у птахів – лише 5-10%. До того ж у деяких організмів увесь геном може існувати в клітинах у вигляді декількох копій. Аналіз ядерного геному птахів доводить, що зменшення розміру їх геному відбулось насамперед завдяки скороченню числа цих некодуючих послідовностей. Тобто птахи на певному етапі свого розвитку пройшли потужний відбір на скорочення розміру геному. Дослідники пов’язують це з необхідністю швидкої генної регуляції та зменшення загального об’єму тіла як пристосування до польоту.
На перший погляд економія на якихось шматках молекул ДНК здається беззмістовною, але треба мати на увазі, що збереження та функціонування ДНК в клітині потребує багато додаткових біохімічних механізмів, які також впливають на розмір клітини та її енергоспоживання. Це, в свою чергу, впливає на швидкість поділу клітини, а через це, на швидкість росту організму (докладніше про те, які наслідки несе збільшення розміру геному, можна прочитати тут - http://elementy.ru/news/432513). Доказом цих закономірностей є вивчення геномів соколів в рамках даної роботи, яке свідчить, що ці надзвичайно активні птахи пройшли ще більш жорсткий відбір на зменшення розміру геному.
Беззубість птахів
Ще однією важливою ознакою птахів є відсутність зубів в щелепах. Ця ознака не дуже розповсюджена серед хребетних тварин:зубів не має у черепах, які втратили зуби значно раніше, ніж птахи, та частково у деяких ссавців (кити, неповнозубі та деякі інші).
Головним питанням беззубості птахів є те, чи втрата зубів відбулась у загального предка всіх сучасних птахів, або у предків різних сучасних груп незалежно. За формування зубів відповідні шість генів, які є спільними для всіх хребетних тварин. Сучасні методи дозволяють встановити, які з цих генів були деактивовані у беззубих форм, та коли приблизно це відбулось.
В процесі порівняння геномів виявилося, що залишки цих генів є в усіх беззубих форм хребетних. У різних груп ссавців до втрати зубів призвели різні мутації, що свідчить про незалежність процесів у різних групах. У всіх сучасних груп птахів залишки генів, що відповідали за формування зубів, мають схожі мутації, що говорить про втрату зубів ще їх далекими спільними предками. Аналіз показав, що інактивація генів, що відповідають за втрату емалі, відбулась приблизно 116 мільйонів років тому, а дентину – 101 мільйон років тому. Одночасно з цим формувались гени, відповідальні за формування рамфотеки (рогового чохла дзьобу). Дослідження свідчили, що втрата зубів починалась з переднього краю щелеп, там саме починав формуватись дзьоб, і після цього процес зсувався назад, охоплюючи всю щелепу.
Цікаво, що серед птахів в широкому сенсі втрата зубів відбувалась неодноразово. Так, в крейдовому періоді поруч з предками сучасних птахів, неорнісами (частина з яких була зубатою, а частина – беззубою), існували енанціорніси, які незалежно втратили зуби, та ще деякі зубаті форми, нащадки яких вимерли наприкінці періоду. Автори доходять висновку, що «у птахів існувала еволюційна схильність до втрати зубів».
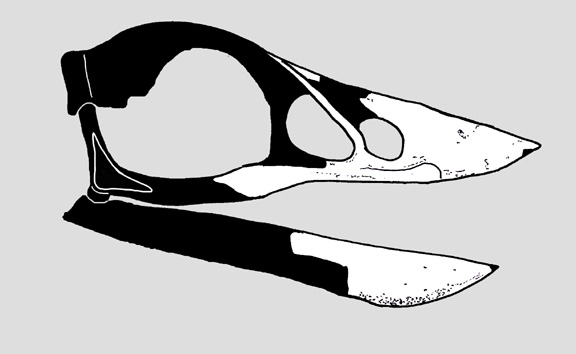
Череп Gobypterix minuta – одного з небагатьох енанціорнісів, який втратив зуби. Мешкав у другій половині крейдового періоду.

Реконструкція Gansus – одного з найбільш давніх представників віялохвостих птахів, що мешкав в крейдовому періоді близько 110 мільйонів років тому. Ці птахи мали зуби.
Що зробило птахів птахами?
Безумовно, поточне дослідження стало важливим етапом у вивченні еволюційної історії птахів. Нові методи призвели до багаторазового збільшення обсягів зібраної інформації та підштовхнули до розробки нових способів аналізу даних. Але говорити про повне з’ясування питань систематики птахів ще зарано. З високим ступенем достовірності були підтверджені деякі останні ідеї щодо загальної еволюції птахів, окреслені загальні риси процесів. Але потрібна ще велика робота по розв’язанню питань еволюції менших за рангом таксонів. Значна частина проаналізованих груп самі по собі є дуже різноманітними, і включення більшого числа видів допоможе краще зрозуміти цей заплутаний ранній період еволюції птахів. Самі автори кажуть, що поступ в основному можливий у частині розробки більш вдосконалених методів аналізу геномних даних та у включенні все більшого числа видів до аналізу.
Дані молекулярних методів все ще мають багато протиріч з більш «традиційною» систематикою. Схоже, що для синтезу даних цих двох підходів потрібен деякий час, коли відбуватиметься накопичення нових матеріалів, розробка ефективних методів їх аналізу та узгодження з результатами інших досліджень.
Загалом можна сказати що відповіді на багато питань еволюції птахів криються в тому короткому та бурхливому періоді в 6 мільйонів років, підчас якого в основному і було закладене те різноманіття, яке ми спостерігаємо зараз.
За матеріалами:
- Зеленков, Н.В. (2013) Система птиц (Aves:Neornithes) в начале ХХІ века. Труды Зоологического института РАН, Приложение 2, 174-190.
- Jarvis, E.D. et al. (2014). Whole-genome analyses resolve early branches in the tree of life of modern birds. Science 346, 1320-1331.
- Meredith, R.W.et al. (2014). Evidence for a single loss of mineralized teeth in the common avian ancestor.Science 346, 1254390.